S-nitrosation
S-nitrosation (also referred to as S-nitrosylation) is a reversible redox-based post-translational modification involving the covalent attachment of a nitric oxide (NO)-derived moiety to reactive cysteine thiols, forming S-nitrosothiols (SNOs) (Pande et al., 2022; Lin et al., 2025; Sedlářová et al., 2025). In plants, it is among the best-characterized NO-dependent modifications and constitutes a central mechanism by which NO exerts its bioactivity as a selective and tightly regulated signaling process rather than a passive by-product (Fancy et al., 2017; Kolbert et al., 2019; Jedelská et al., 2020; Treffon et al., 2021; Liao et al., 2023; Lin et al., 2025). S-nitrosation operates as a reversible molecular switch, analogous to phosphorylation, that modulates protein activity, conformation, stability, localization, and interactions, thereby controlling key physiological processes including germination, root development, stomatal function, and stress responses (Corpas and Palma, 2018; Liu et al., 2024; Kolbert et al., 2024; Lin et al., 2025). Its specificity is determined by the local redox environment, cysteine reactivity, and the dynamic interplay between nitrosation, transnitrosation, and denitrosation reactions (Jedelská et al., 2020; Treffon et al., 2021; Lin et al., 2025; Das et al., 2025).
S-nitrosoglutathione (GSNO), generated from NO and glutathione (GSH), functions as a principal intracellular reservoir and mobile carrier of NO bioactivity, enabling spatial propagation of NO signals within and between plant cells (Gupta et al., 2011; Liao et al., 2023; Liu et al., 2024). Cellular S-nitrosation levels are tightly controlled by S-nitrosoglutathione reductase (GSNOR), which regulates GSNO turnover and thereby indirectly modulates protein S-nitrosation (Lin et al., 2025). This modification occurs across major cellular compartments, including the cytosol, nucleus, chloroplasts, and mitochondria, underscoring its broad regulatory scope (Liao et al., 2023; Lin et al., 2025).
Mechanisms and Regulation
S-nitrosation arises through multiple, context-dependent biochemical pathways governed by the redox environment and the availability of reactive nitrogen species (RNS) (Lin et al., 2025). Direct nitrosation is primarily mediated by NO-derived oxidized intermediates, such as dinitrogen trioxide (N2O3), which efficiently target cysteine residues under aerobic conditions (Wei et al., 2024; Lin et al., 2025). In vivo, transnitrosation constitutes a dominant mechanism, whereby NO groups are transferred from low-molecular-weight S-nitrosothiols, particularly GSNO, or from S-nitrosated proteins to specific cysteine residues, enabling amplification and propagation of NO signaling (Wei et al., 2024; Liu et al., 2024). Transition metals, including iron and copper, can further facilitate localized nitrosation reactions (Lin et al., 2025). Site selectivity is dictated by cysteine physicochemical properties -such as thiol pKa, solvent accessibility, and the surrounding amino acid environment- ensuring that S-nitrosation targets defined protein subsets rather than occurring indiscriminately (Pande et al., 2022; Lin et al., 2025). S-nitrosation is dynamically reversible. Thioredoxins (Trx) directly catalyze denitrosation of specific protein targets, whereas GSNOR regulates SNO homeostasis indirectly through GSNO degradation (Jedelská et al., 2020; Treffon et al., 2021; Liao et al., 2023; Lin et al., 2025). In addition, NADPH-dependent GSNO catabolism mediated by aldo-keto reductases (AKRs) introduces an additional regulatory layer (Jedelská et al., 2020; Treffon et al., 2021; Das et al., 2025). Together, these systems establish tight spatiotemporal control over NO signaling.
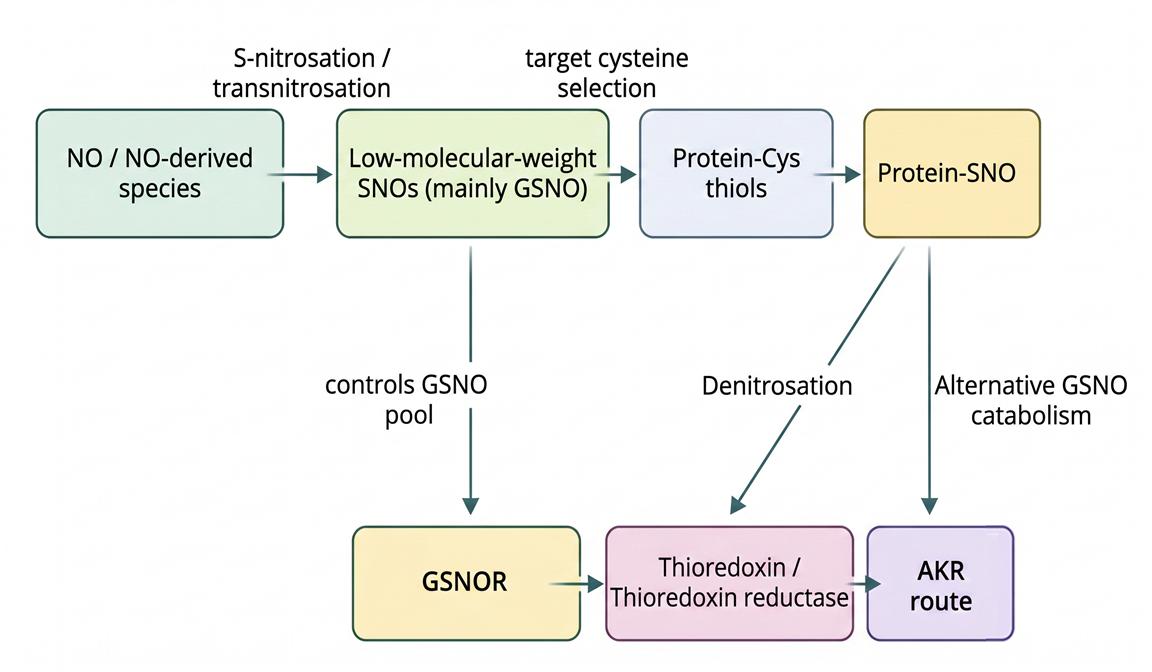
Fig. 1. Mechanisms and regulation of protein S-nitrosation in plants. NO and NO-derived reactive species promote S-nitrosation either directly or via transnitrosation through low-molecular-weight S-nitrosothiols, primarily GSNO. Target specificity is determined by the local protein environment and cysteine reactivity. The S-nitrosation status is dynamically regulated by denitrosation systems, including Trx/thioredoxin reductase and GSNOR, as well as alternative GSNO catabolic pathways such as aldo-keto reductases (AKRs). This balance ensures reversible and selective NO-dependent signaling. Adapted from: Lin et al., 2025; Liao et al., 2023; Wei et al., 2024; Liu et al., 2024; Das et al., 2025.
Biological Functions and Redox Integration
S-nitrosation is integral to the regulation of plant growth, development, and signaling networks through its effects on enzymes, transcription factors, and signaling proteins (Liu et al., 2024; Lin et al., 2025). It governs processes such as germination, root development, flowering, and senescence, and modulates gene expression by altering transcription factor activity and DNA-binding capacity (Liao et al., 2023; Wei et al., 2024). At the systems level, S-nitrosation operates at the interface of hormone signaling, redox regulation, and stress adaptation. It contributes to auxin and abscisic acid (ABA) pathways and modulates defense responses through NO/ROS crosstalk (Kolbert et al., 2019; Pande et al., 2022; Sedlářová et al., 2025). Moreover, it interacts with other post-translational modifications, including phosphorylation, tyrosine nitration, and glutathionylation, forming interconnected regulatory networks. The balance between S-nitrosation and denitrosation defines cellular redox homeostasis. GSNO functions as a central NO reservoir, while GSNOR and thioredoxin systems regulate its turnover and protein SNO levels (Jedelská et al., 2020; Treffon et al., 2021; Das et al., 2025). Disruption of this balance leads to nitrosative stress and impaired cellular function, whereas coordinated interactions with reactive oxygen species (ROS) enable fine-tuned signaling responses (Wei et al., 2024; Liu et al., 2024).
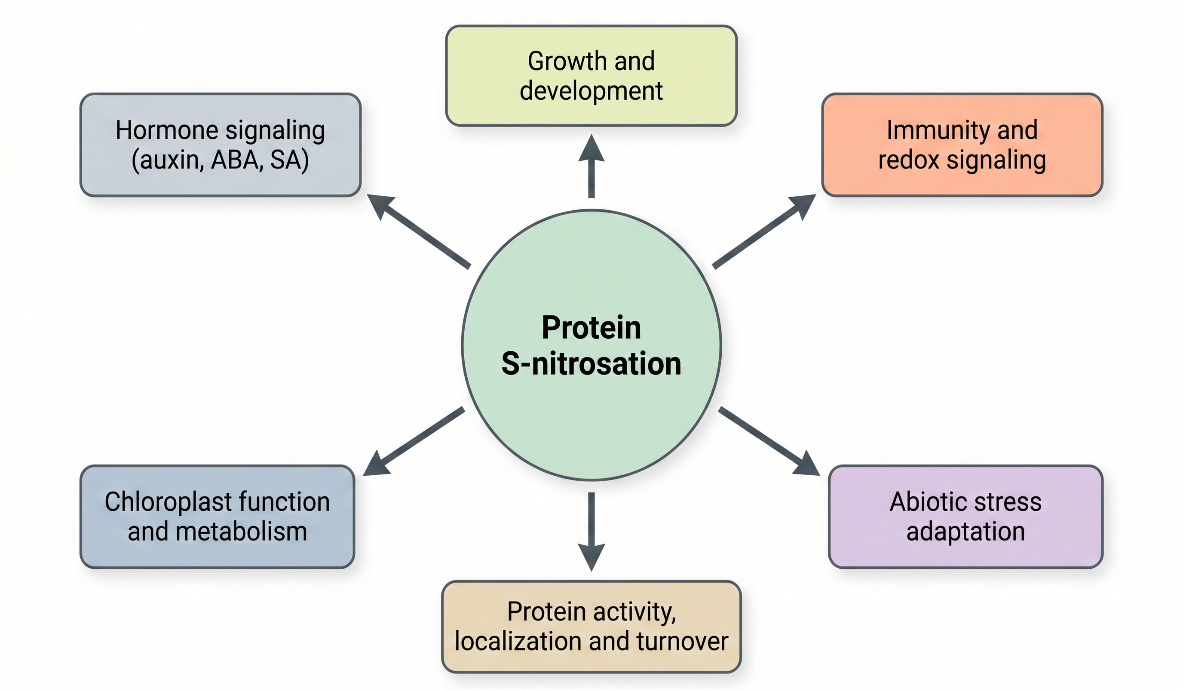
Fig. 2. Functional roles of protein S-nitrosation in plant physiology. S-nitrosation acts as a central regulatory mechanism influencing multiple biological processes, including growth and development, hormone signaling (auxin, abscisic acid, salicylic acid), chloroplast function and metabolism, immune responses, and adaptation to abiotic stress. By modulating protein activity, localization, and turnover, S-nitrosation integrates nitric oxide signaling with redox homeostasis and plant stress responses. Adapted from: Liao et al., 2023; Liu et al., 2024; Wei et al., 2024; Lin et al., 2025; Sedlářová et al., 2025.
Role in Stress Responses
S-nitrosation is a key mediator of plant responses to abiotic and biotic stresses, including drought, salinity, heavy metal exposure, and pathogen attack (Kolbert et al., 2019; Liao et al., 2023; Lin et al., 2025). It regulates stress-responsive proteins, particularly antioxidant and redox-related enzymes, thereby maintaining cellular homeostasis (Wei et al., 2024; Liu et al., 2024). Depending on the target, S-nitrosation can enhance or suppress protein activity, enabling flexible adaptation to environmental challenges (Liu et al., 2024). Emerging evidence further links environmental factors, including metal exposure and nanoparticle interactions, to S-nitrosation dynamics, connecting NO signaling with stress adaptation at the nano-bio interface.
Detection and Analytical Approaches
Analytical strategies for S-nitrosation have progressed from the classical biotin-switch technique (BST) to advanced chemistries such as FAT-switch and high-throughput LC-MS/MS-based proteomics (Pande et al., 2022; Lin et al., 2025; Wei et al., 2024). These approaches have substantially expanded the known plant S-nitrosoproteome. However, the labile and reversible nature of S-nitrosation complicates the discrimination of physiologically relevant modifications from non-specific or stress-induced events, necessitating rigorous validation (Pande et al., 2022; Lin et al., 2025).
References
- Corpas, F. J., & Palma, J. M. Nitric oxide on/off in fruit ripening. Plant Biology 20(5), 805-807 (2018). https://doi.org/10.1111/plb.12852
- Das, A. K., Lee, D. S., Lee, G. J., Kim, Y. S., Hussain, S., Lee, M. S., ... & Mun, B. G. The Central Role of GSNOR: Decoding Nitric Oxide Signaling for Crop Stress Tolerance. International Journal of Molecular Sciences 26(23), 11486 (2025). https://doi.org/10.3390/ijms262311486
- Fancy, N. N., Bahlmann, A. K., & Loake, G. J. Nitric oxide function in plant abiotic stress. Plant, Cell & Environment 40(4), 462-472 (2017). https://doi.org/10.1111/pce.12707
- Gupta, K. J., Fernie, A. R., Kaiser, W. M., & van Dongen, J. T. On the origins of nitric oxide. Trends in Plant Science 16(3), 160-168 (2011). https://doi.org/10.1016/j.tplants.2010.11.007
- Jedelská, T., Luhová, L., & Petřivalský, M. Thioredoxins: emerging players in the regulation of protein S-nitrosation in plants. Plants 9(11), 1426 (2020). https://doi.org/10.3390/plants9111426
- Kolbert, Z. S., Barroso, J. B., Brouquisse, R., Corpas, F. J., Gupta, K. J., Lindermayr, C., ... & Hancock, J. T. A forty year journey: The generation and roles of NO in plants. Nitric Oxide 93, 53-70 (2019). https://doi.org/10.1016/j.niox.2019.09.006
- Kolbert, Z., Barroso, J. B., Boscari, A., Corpas, F. J., Gupta, K. J., Hancock, J. T., ... & Loake, G. J. Interorgan, intraorgan and interplant communication mediated by nitric oxide and related species. New Phytologist 244(3), 786-797 (2024). https://doi.org/10.1111/nph.20085
- Liao, W., Igamberdiev, A. U., & Palma, J. M. Advances in nitric oxide signalling and metabolism in plants. International Journal of Molecular Sciences 24(7), 6397 (2023). https://doi.org/10.3390/ijms24076397
- Lin, W., Shang, J. X., Li, X. Y., Zhou, X. F., & Zhao, L. Q. Nitric oxide regulates multiple signal pathways in plants via protein S-nitrosylation. Current Issues in Molecular Biology 47(6), 407 (2025). https://doi.org/10.3390/cimb47060407
- Liu, Y., Liu, Z., Wu, X., Fang, H., Huang, D., Pan, X., & Liao, W. Role of protein S-nitrosylation in plant growth and development. Plant Cell Reports 43(8), 204 (2024). https://doi.org/10.1007/s00299-024-03290-z
- Pande, A., Mun, B. G., Rahim, W., Khan, M., Lee, D. S., Lee, G. M., ... & Yun, B. W. Phytohormonal regulation through protein s-nitrosylation under stress. Frontiers in Plant Science 13, 865542 (2022). https://doi.org/10.3389/fpls.2022.865542
- Sedlářová, M., Jedelská, T., Lebeda, A., & Petřivalský, M. Progress in plant nitric oxide studies: implications for phytopathology and plant protection. International Journal of Molecular Sciences 26(5), 2087 (2025). https://doi.org/10.3390/ijms26052087
- Treffon, P., Rossi, J., Gabellini, G., Trost, P., Zaffagnini, M., & Vierling, E. Quantitative proteome profiling of a S-nitrosoglutathione reductase (GSNOR) null mutant reveals a new class of enzymes involved in nitric oxide homeostasis in plants. Frontiers in Plant Science 12, 787435 (2021). https://doi.org/10.3389/fpls.2021.787435
- Wei, L., Liao, W., Zhong, Y., Tian, Y., Wei, S., & Liu, Y. NO-mediated protein S-nitrosylation under salt stress: Role and mechanism. Plant Science 338, 111927 (2024). https://doi.org/10.1016/j.plantsci.2023.111927